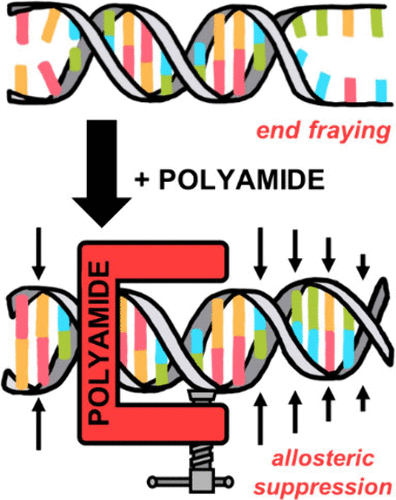
Long-Range Allosteric Modulation of DNA Duplex Dynamics Induced by Pyrrole-Imidazole Polyamide Binding Read More »
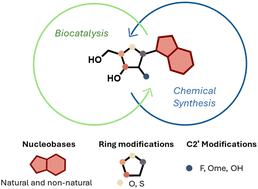
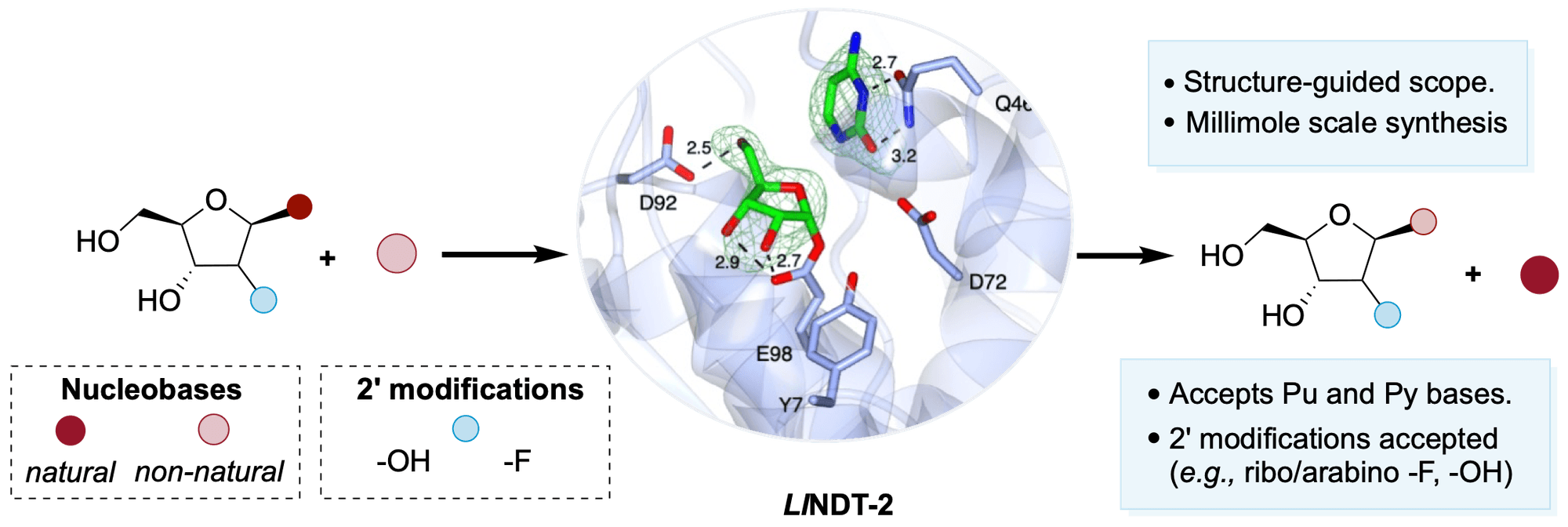
Preparation of nucleoside analogues: opportunities for innovation at the interface of synthetic chemistry and biocatalysis Read More »
Factors to Consider for Synthesis in 1536-Well Plates─An Amide Coupling Case Study for PROTAC Synthesis Read More »
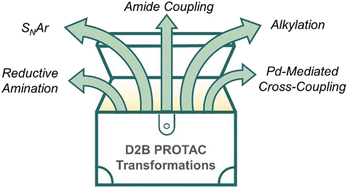
Expanding the reaction toolbox for nanoscale direct-to-biology PROTAC synthesis and biological evaluation Read More »
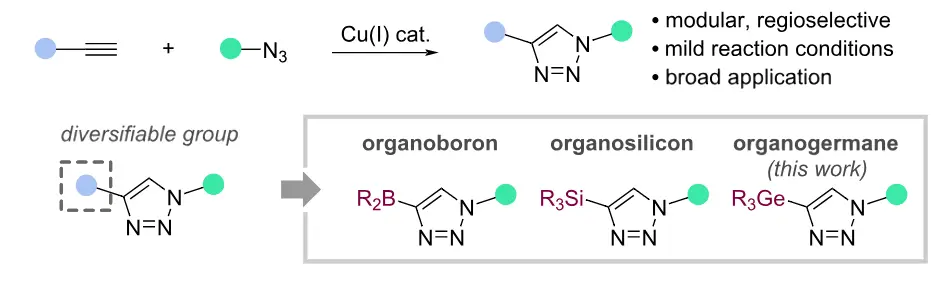
Germanyl triazoles as a platform for CuAAC diversification and chemoselective orthogonal cross-coupling Read More »
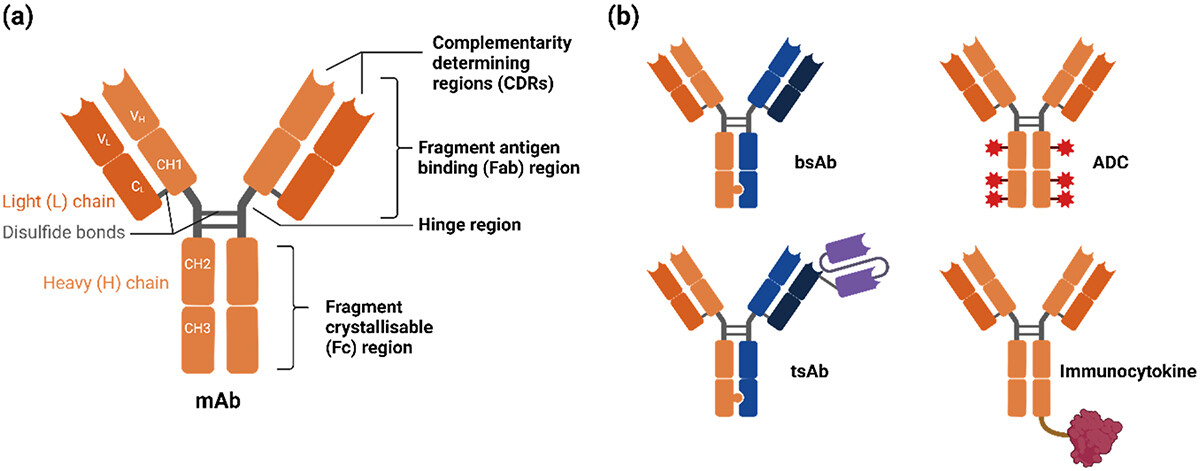
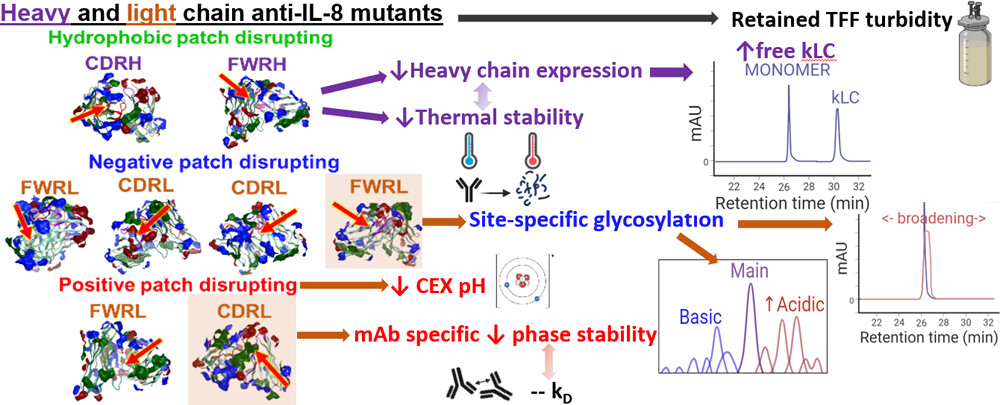
Assessing the Manufacturability and Critical Quality Attribute Profiles of Anti-IL-8 Immunoglobulin G Mutant Variants Read More »
Core Modifications of GSK3335103 toward Orally Bioavailable αvβ6 Inhibitors with Improved Synthetic Tractability Read More »
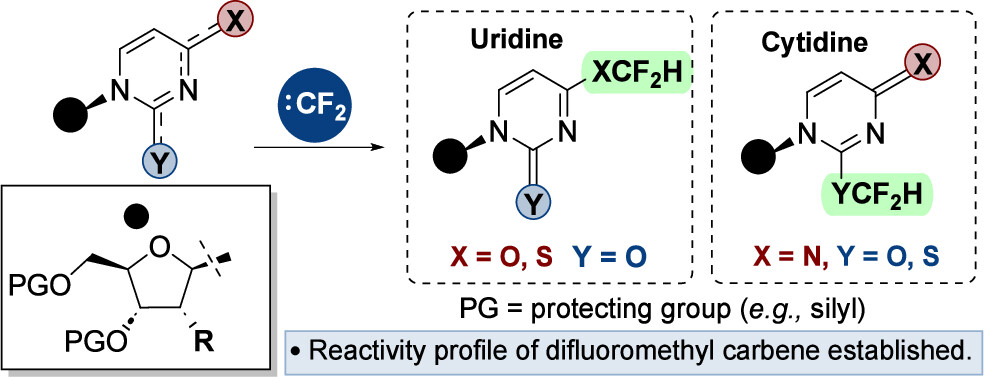
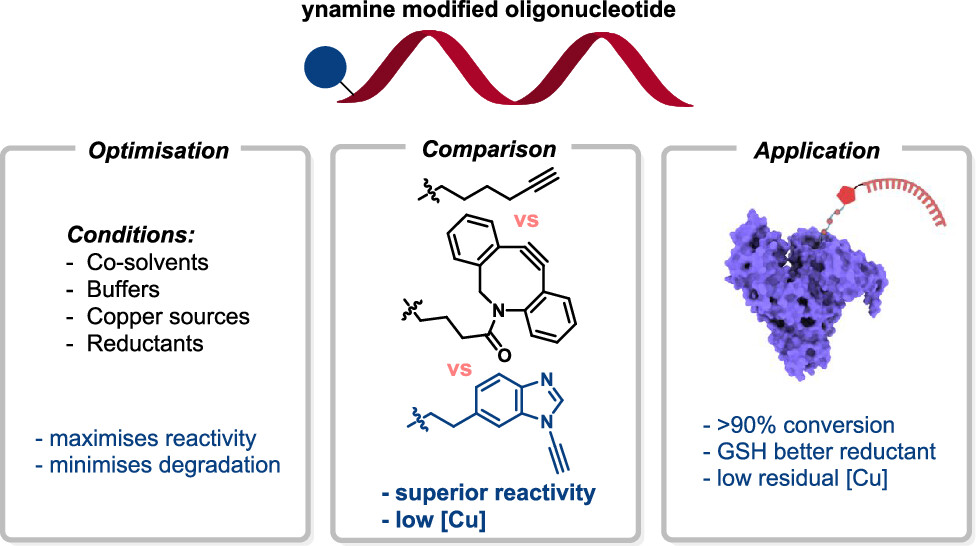
Reactivity Profiling for High-Yielding Ynamine-Tagged Oligonucleotide Click Chemistry Bioconjugations Read More »